SA JOURNAL OF DIABETES & VASCULAR DISEASE
REVIEW
VOLUME 11 NUMBER 2 • JUNE 2014
77
Nitric oxide (NO) is the main vasodilator from the endothelium
specifically involved in blood flow and blood distribution, and
while reduction in nitric oxide synthesis lowered total blood flow,
exercise-mediated capillary recruitment was not affected.
46
In fact,
inhibition of NO formation enhances both resting and exercise-
mediated muscle oxygen uptake;
47
despite a reduction in total
flow, microvascular flow was not affected, suggesting that NO is
not involved in the vascular response to exercise. However, other
studies have shown that exercise training required nitric oxide for
improvement in flow-induced dilation.
44
It is therefore possible that
while NO is not involved in the acute response to exercise, exercise
training restores general endothelial health, as evidenced by a
restored endothelium-dependent vasodilation in response to flow.
Therefore, as well as the acute effects of exercise, which may be
independent of NO, an exercise regimen may improve endothelial
function.
Metabolic effects of exercise
The distribution of blood through muscle increases the capacity for
nutrient exchange. In exercise, the primary purpose of functional
hyperaemia is for oxygen delivery, as the oxygen required by
exercising muscle is much higher than resting muscle.
37
Recruitment
of capillaries can decrease the velocity of blood flow by increasing
the cross-sectional area of the capillary bed and the time available
for exchange. Recruitment also increases surface area for exchange
and decreases perfusion distances to promote oxygen delivery to
tissues with exercise
34
(Fig. 2). While in exercise the main metabolite
required at the working muscle is oxygen, distribution of other
nutrients can also be affected, including glucose, fats, other
hormones and cytokines. Muscle metabolism can therefore be
altered by perfusion of the tissue.
48,49
While there can be regulated
transport of certain larger hormones across the vasculature,
50,51
smaller molecules can diffuse across the endothelium easily, possibly
making muscle perfusion a more important player in the delivery of
glucose and oxygen to the tissue.
Skeletal muscle is the main site of basal glucose uptake, and
is the tissue most associated with exercise; therefore the effect
of exercise would likely be localised in muscle. A single bout of
exercise in sedentary men increases glucose uptake and glucose
effectiveness, and it was suggested that the increased blood flow
and distribution enhanced glucose delivery to the tissue.
52
Capillary
recruitment with exercise contributes to glucose uptake, but NO is
not required for exercise-mediated capillary recruitment.
46
Instead,
NO augments glucose uptake in high-intensity exercise,
46
but not
low-intensity exercise, and may be involved in a partitioning of fuel
utilisation.
53
Longer-term, mild exercise training improves glucose
disposal, even with no change in body composition.
54
This sustained
effect was independent of the metabolic benefits of a single
bout of exercise. Changes in insulin-specific glucose transporter
expression have been detected after exercise training,
55,56
as have
changes in DNA methylation,
57
but it is also possible that general
improvements in endothelial function increase delivery, and thus
metabolism, of glucose.
Fat deposition in muscle is often thought to be associated with
insulin resistance,
58,59
and selective reduction of intramyocellular
lipid restores normal insulin signalling, reverting to a healthy
metabolic state.
60
Therefore, rather than intramuscular or total
body fat, intramyocellular fat is related to muscle insulin resistance.
However a paradox is noted when athletes are considered, as they
often have very high levels of intramyocellular lipid, yet high insulin
sensitivity.
61
Intramyocellular lipid content is increased after exercise
intervention and diet change, coinciding with an increase in insulin
sensitivity, suggesting that intramyocellular lipid content may not
directly impair cellular insulin sensitivity.
62
Exercise can prevent lipid-
induced insulin resistance,
63
and the form the lipid is stored in may
contribute to insulin resistance, asceramide or diacylglycerol
64-66
are more detrimental to cellular insulin action than triglycerides.
Another component of the divergent effects of intramyocellular
lipid on insulin action could be the site of storage of excess fat. Lipid
droplets within the muscle cell may regulate insulin action
67
and
possibly the mitochondria, such that lipid droplet-derived fats can
be used as fuel by exercising muscle.
68
By contrast, nutrient overload
can alter the lipid droplet coat proteins and change the interaction
of the lipid droplet with other organelles, causing inflammation and
oxidative stress. Therefore, while fat deposition in muscle may not
directly affect vascular function, the resulting inflammation
69
and
oxidative stress
70
from intramyocellular lipid can lead to endothelial
dysfunction. Further, fat deposition in endothelial cells has not been
directly measured, and may occur in a similar fashion as in muscle
and directly affect vascular function.
Muscle is composed of oxidative and glycolytic fibre types, with
oxidative fibres typically having more mitochondria, and being
actively recruited during exercise. The density of capillaries is
greater in oxidative muscle; reduced oxidative activity in type 2
diabetes patients is most likely due to a reduction in slow oxidative
fibres.
71
The decrease in oxidative activity and increase in glycolytic
activity in these patients was closely linked to the fraction of each
fibre type present in muscle, suggesting that type 2 diabetes
patients show both changes in fibre composition and fibre-specific
metabolism. Mitochondrial dysfunction has been proposed to
be both a cause
72
and a consequence
73
of insulin resistance, and
may contribute to endothelial dysfunction.
74
If oxygen delivery is a
component of mitochondrial health and biogenesis, it is possible
that impaired perfusion may contribute to fibre type switching,
where an oxidative fibre, which is typically highly vascularised
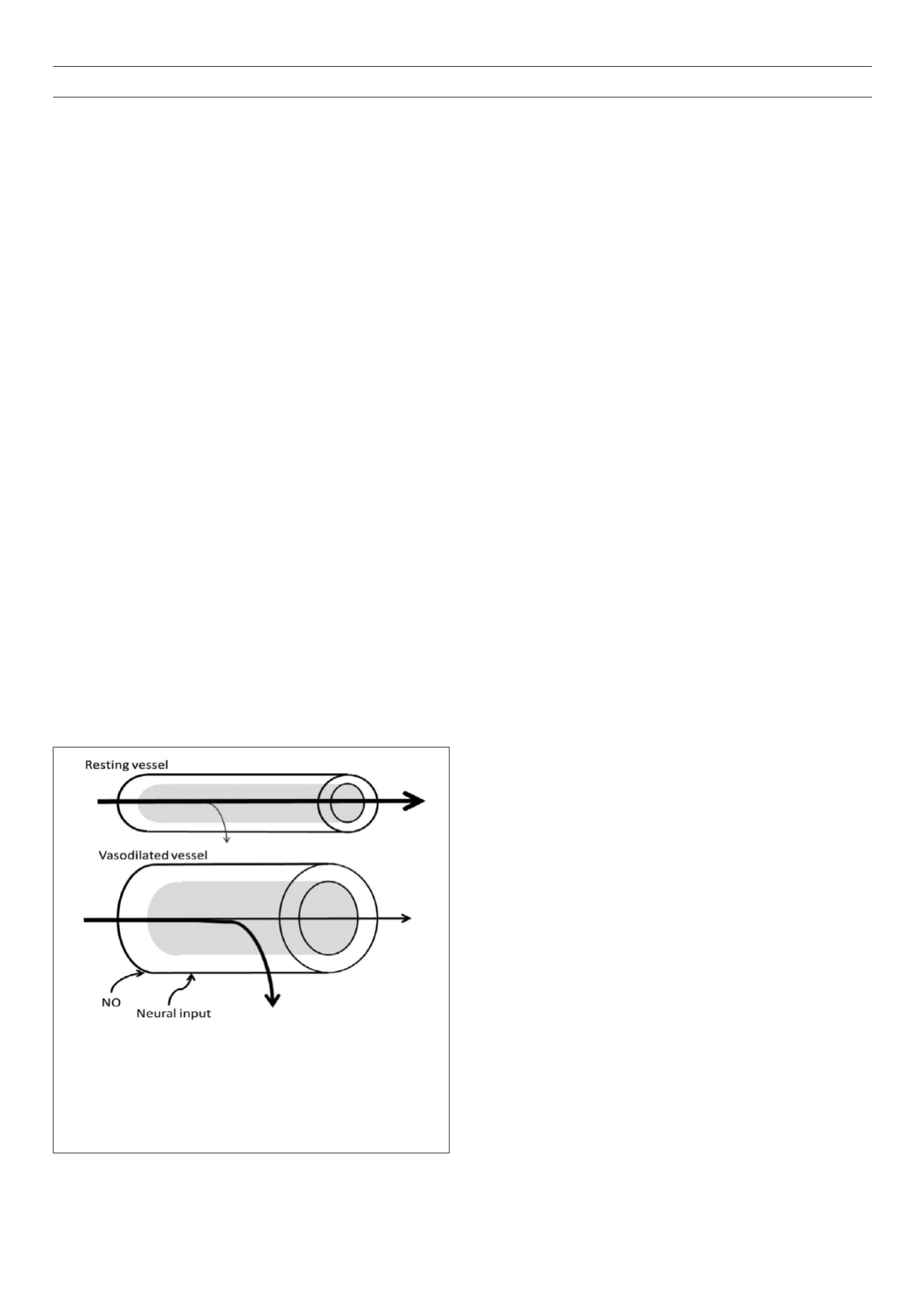
Figure 2.
Vasodilation affects delivery, and therefore metabolism. The rate
of transfer across the endothelium is dependent on surface area, perme-
ability of the endothelium, diffusion distance, and concentration difference
(Fick’s first law of diffusion). Vasodilation increases surface area in arterioles
for exchange, but will also recruit downstream capillaries, which will reduce
diffusion distance and increase surface area for exchange. Working muscle
increases oxygen utilisation, increasing the concentration difference from the
blood vessel to the tissue.


